



An In-Depth Look At Bloom

Figure 1: Pollen grains suspended on the hairs of a painters brush. Apple producers rely on pollinating insects to transfer pollen to the stigmatic surface of blossoms. (Photo courtesy of Tom Kon, Penn State University)
Most fruit growers are familiar with the general concepts of pollination. Pollinating insects seek sugar-rich nectar, or protein-rich pollen for food. Apple trees lure these insects using visual cues (petals), chemical attractants (fragrance), nectar, and pollen.
While the pollinators visit flowers to collect food resources, apple blossoms use pollinator visitation as an opportunity to capture pollen and procreate.
After apple pollen (Figure 1) is deposited on the stigmatic surface of an apple pistil (female portion of the flower, see Figure 2), the romantic notion that accompanies the plant-pollinator interaction suddenly disappears and is replaced with a series of complex biochemical interactions between pollen and pistil. Pollen tube growth of apple has been studied for more than 100 years, and we still have more to learn. So how does it all work?

Figure 2: The pistil (female portion of the flower) is composed of the stigma, style, and ovary. Pollen is received and hydrated by the stigma, resulting in germination and growth in the style. The style supports and directs pollen tube growth to the ovary, which contains the ovules. If successful, pollen tubes penetrate the ovules and rupture, resulting in fertilization. (Photo courtesy of Tom Kon, Penn State University)





Pollen Germination And Growth
In the process of gathering food resources, pollinating insects come in contact with the stigmatic surface (Figure 2) and deposit pollen. The stigmatic surface of an apple blossom is comprised of globular cells that begin to break down as soon as the blossom opens. As these cells break down they emit a sticky secretion, primarily composed of carbohydrates and proteins, which aids in pollen adhesion and germination. Pollen grains adhere to the stigmatic surface, hydrate, and germinate. After penetrating the stigma, pollen tubes begin growth toward the ovaries in the intercellular spaces of the style (Figure 3). Pollen tubes grow exclusively at the tip, and under optimal conditions, can travel the length of the style in 2 or 3 days.
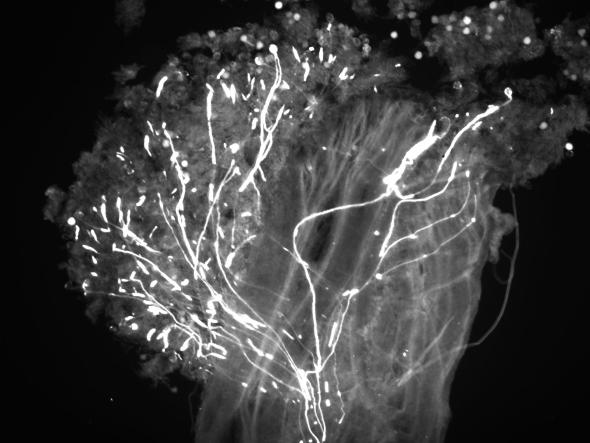
Figure 3: Pollen hydration, germination, and growth on the stigmatic surface and upper style. When stained and observed via fluorescence microscopy, pollen tubes glow vividly. After pollen is deposited on the stigma, the course and rate of pollen tube growth is dictated by biochemical signals, genetics, and environmental conditions. (Photo courtesy of Tom Kon, Penn State University)
During germination and early growth in the style, pollen tubes primarily use stored energy from the pollen grain. After these reserves are exhausted, the pollen tube relies on nutrient resources provided by the style. Pollen tube growth is directed by resources and signals emitted by style and ovary tissues.
Recent evidence suggests that resources are limited in the lower style when compared to the upper style, prompting competition between pollen tubes. These resources/signals guide pollen tubes through the style, eventually leading to the ovary. If successful, pollen tubes reach and fertilize the ovule(s).
Individual apple blossoms don’t open at the same time in a blossom cluster: king blossoms open first and lateral blossoms open later. This extends the opportunity for pollinator visitation, and increases the chances of pollination and fruit set at each spur. Flower position within blossom clusters may also influence the duration of stigmatic receptivity, and the ability for pollen grains to adhere to the stigmatic surface.
Recent evidence suggests that king blossoms are receptive for a shorter period of time when compared to lateral blossoms, however, greater numbers of pollen grains adhered to and germinated on stigmas of king blossoms. Maximum receptivity for king blossoms was observed at or shortly after opening, while receptivity for lateral blossoms was slightly delayed.
If this process wasn’t already complicated enough, consider this: female flower tissues can begin to degrade as soon as the blossom opens. Temperature is the primary factor that influences the rate of flower degradation. High temperatures increase the rate of pollen tube growth, but hasten the degradation of stigmatic and ovule receptivity. Conversely, cool temperatures reduce the rate of pollen tube growth, but the duration of stigmatic and ovule receptivity is increased. Extremely hot or cold temperatures during bloom can be detrimental to fruit set.
The number of days that a blossom can potentially produce a fruit is referred to as the Effective Pollination Period (EPP). EPP is determined by the rate of ovule and stigma degeneration, due to temperature. Insect activity is also influenced by temperature, so warm temperatures increase pollinator activity while cool temperatures reduce activity.

(Photo courtesy of Tom Kon, Penn State University)
Additional Factors
There are a few factors that influence pollen tube growth and floral receptivity. These include: variety, location, flower quality, branch angle, and tree branch angle.
Variety: The variety of both female (pistil) and male (pollen) can influence the rate of pollen tube growth. Researchers at Virginia Tech in conjunction with the Washington Tree Fruit Research Commission have created models that estimate rate of pollen tube growth in several commercially important apple varieties. Since pollen tube growth rate is influenced by the pistil, specific models had to be developed for each variety. These models are of practical value, and are used as a timing aid for blossom thinner applications.
Researchers at Washington State University will begin screening crabapples with the goal of finding improved pollinizers with rapid pollen tube growth rates, disease resistance, high pollen viability, full compatibility, annual flower production, and bloom periods that coincide with commercial apple varieties.
Location: Aside from environmental differences due to location, there appears to be variation in the average style length of the same cultivar at different sites. Differences were also observed in style length of strains of the same variety. Simply put, longer styles require pollen tubes to travel a greater distance to fertilize the ovules. It is unclear, however, if the length of apple styles can have an influence on the productivity of orchard sites.
Flower Quality: Flower quality appears to play a role in the longevity of apple blossoms, and fruit set potential. It is difficult to define what constitutes a “high quality” flower since flowers are produced in a mixed bud, containing spur leaves and vegetative shoots. Flower quality is likely influenced by several factors, including: variety, rootstock, nutrition, cropping status, branch angle, and tree/branch age. Nitrogen and boron status play a role in blossom longevity and pollen tube growth, respectively. Cropping status in the previous year can influence flower structure and subsequent fruit set. While still functional, flowers from heavily cropped trees have a shorter EPP.
Branch Angle: Blossoms from upright limbs have a lower potential to set fruit when compared to limbs in a horizontal position. Branch angle does not influence the physical structure of blossoms, but the duration of ovule viability is shorter in upright limbs.
Tree And Branch Age: In general, blossoms from young wood and trees contain ovules of poor quality. There are appear to be exceptions to this rule, as some varieties appear to have no problems setting fruit on young wood (Gala).
How To Ensure Pollination Is Not A Problem
Apple producers rely on plant-pollinator interactions to increase the chances that fruit set is not a limiting factor in production. To assure that pollination is not a limiting factor in production; orchardists increase pollinator insect density, plant compatible pollinizer varieties with a similar bloom window within or in adjacent blocks, and eliminate flowering weeds that may attract pollinators. After pollen is deposited on the stigma, the course of pollen tube growth is dictated by chemical signals, genetics, and environmental conditions. Apple trees have reproductive strategies to increase the opportunity for successful fertilization and fruit set, but temperature is a key factor in reproductive success and Mother Nature can be a cruel partner.
While pollination is a self-regulated process, management decisions can influence the viability of apple blossoms. Existing recommendations for crop load management, plant nutrition, and tree training can improve flower quality and the potential for initial fruit set. While pollinators certainly play a key role in production, the story is just beginning after the bees fly away. The continued study of the apple reproductive system – and our manipulation of it – is important.
Additional information about apple pollination can be found online here.
Subscribe Today For
